
Six Novel Mycoviruses Containing Positive Single-Stranded RNA and Double-Stranded RNA Genomes Co-Infect a Single Strain of the Rhizoctonia solani AG-3 PT

Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction and Purification of RNA
2.2. Metatranscriptome Sequencing
2.3. Validation of Mycoviruses in Strain ZJ-2H
2.4. Determination of Full Length cDNAs of Putative Mycoviruses
2.5. Sequence Analysis
2.6. Extraction and Observation of Virus Particles
2.7. Confirmation of the Mycoviruses That Assemble into Virus Particles
3. Results
3.1. Identification of Mycoviruses

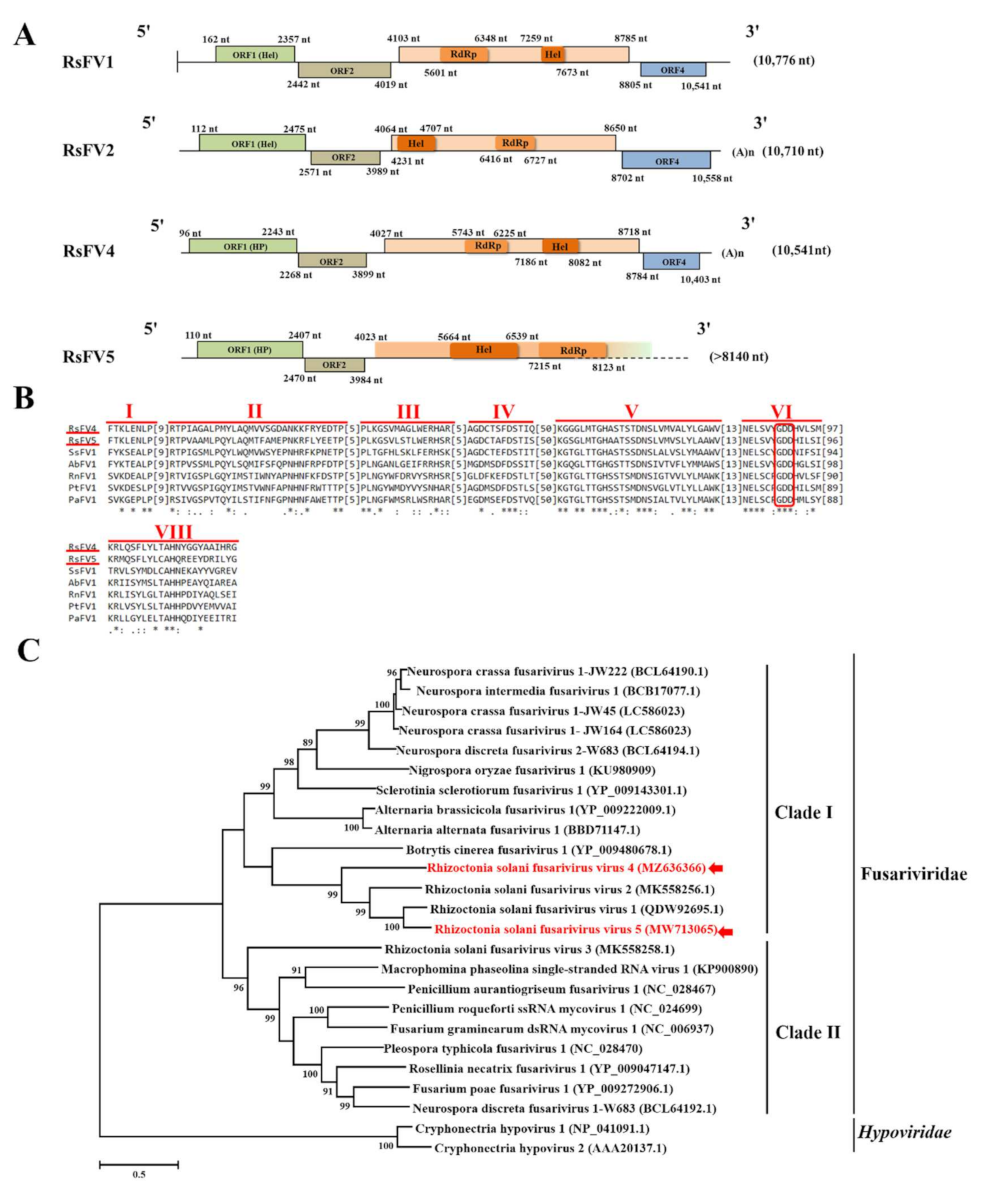
3.2. Genome Organization and Phylogenetic Analysis of Putative Members of the Proposed Family Fusariviridae

3.3. Genome Organization and Phylogenetic Analysis of the Putative Member of the Family Mitoviridae

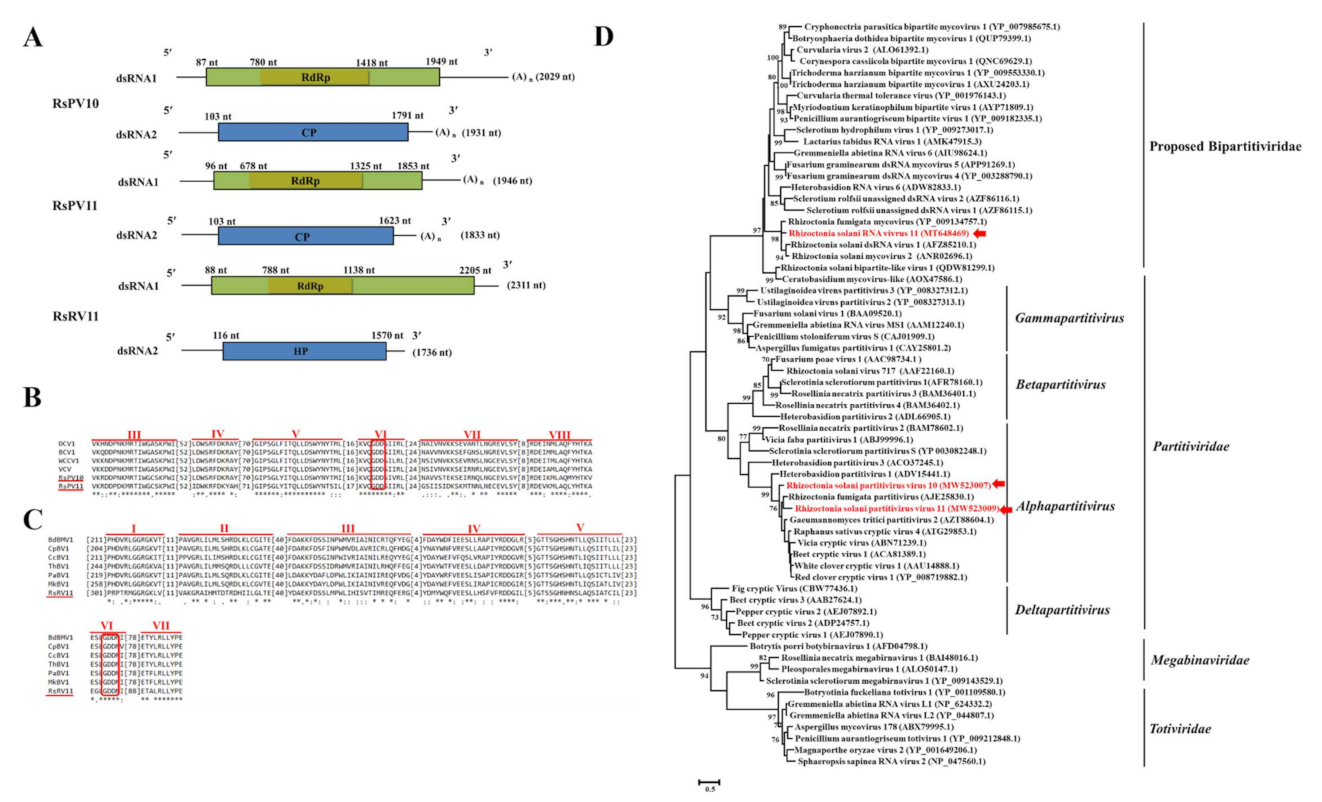
3.4. Genome Organization and Phylogenetic Analysis of Partiti-Like Viruses

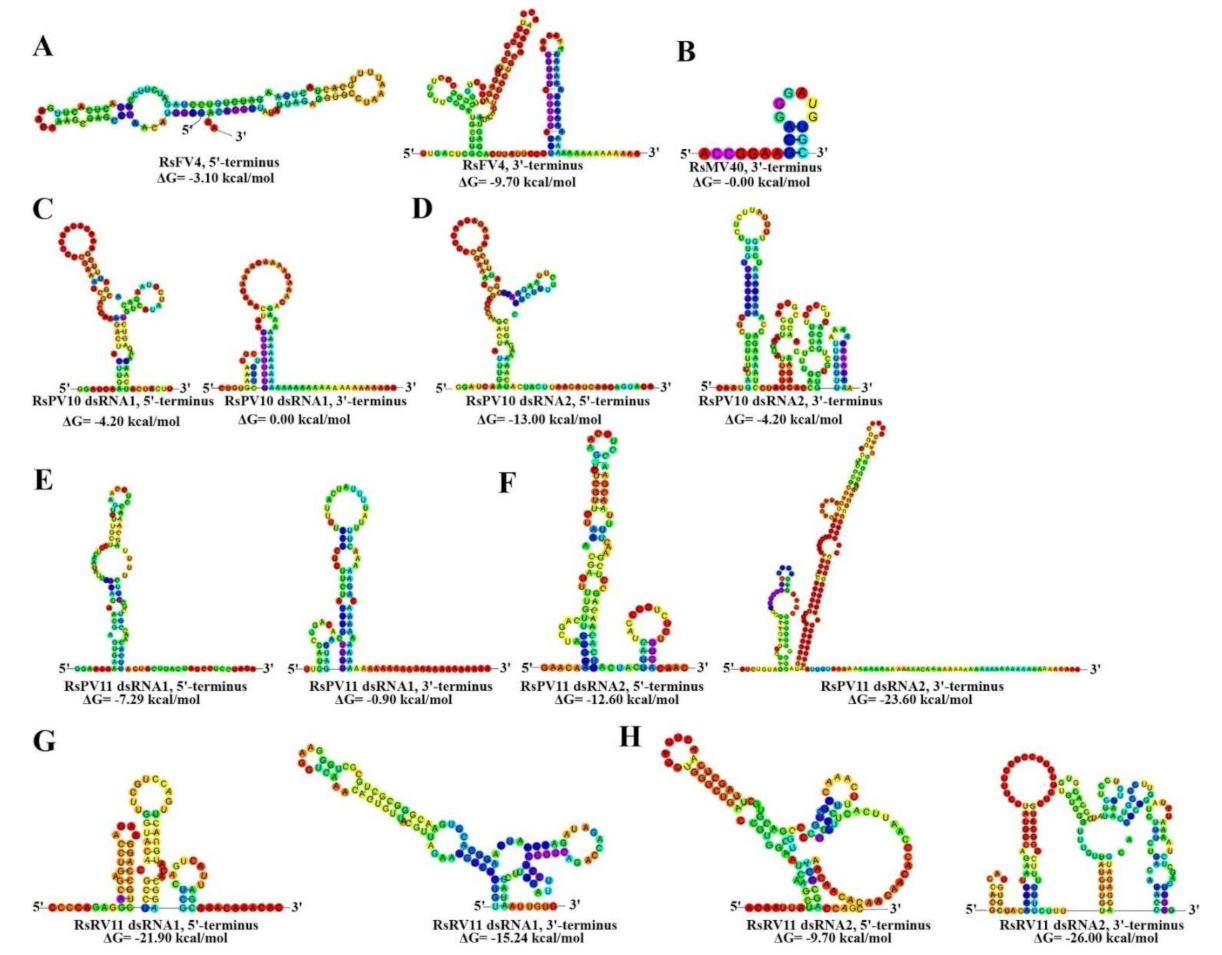
3.5. Analysis of the Secondary Structures of the Termini of Mycoviruses

3.6. Observations of Virus Particles and Confirmation of the Mycoviruses Contributing to the Assembly of Virus Particles
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kotta-Loizou, I. Mycoviruses: Past, present, and future. Viruses 2019, 11, 361. [Google Scholar] [CrossRef] [Green Version]
- Myers, J.M.; Bonds, A.E.; Clemons, R.A.; Thapa, N.A.; Simmons, D.R.; Carter-House, D.; Ortanez, J.; Liu, P.; Miralles-Duran, A.; Desiro, A.; et al. Survey of early-diverging lineages of fungi reveals abundant and diverse mycoviruses. mBio 2020, 11, e02027-20. [Google Scholar] [CrossRef] [PubMed]
- Bormann, J.; Heinze, C.; Blum, C.; Mentges, M.; Brockmann, A.; Alder, A.; Landt, S.K.; Josephson, B.; Indenbirken, D.; Spohn, M.; et al. Expression of a structural protein of the mycovirus FgV-ch9 negatively affects the transcript level of a novel symptom alleviation factor and causes virus infection-like symptoms in Fusarium graminearum. J. Virol. 2018, 92, e00326-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Marzano, S.Y.L.; Nelson, B.D.; Ajayi-Oyetunde, O.; Bradley, C.A.; Hughes, T.J.; Hartman, G.L.; Eastburn, D.M.; Domier, L.L. Identification of diverse mycoviruses through metatranscriptomics characterization of the viromes of five major fungal plant pathogens. J. Virol. 2016, 90, 6846–6863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osaki, H.; Sasaki, A.; Nomiyama, K.; Tomioka, K. Multiple virus infection in a single strain of Fusarium poae shown by deep sequencing. Virus Genes 2016, 52, 835–847. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–550. [Google Scholar] [CrossRef]
- Dolja, V.V.; Koonin, E.V. Metagenomics reshapes the concepts of RNA virus evolution by revealing extensive horizontal virus transfer. Virus Res. 2018, 244, 36–52. [Google Scholar] [CrossRef]
- Gilbert, K.B.; Holcomb, E.E.; Allscheid, R.L.; Carrington, J.C. Hiding in plain sight: New virus genomes discovered via a systematic analysis of fungal public transcriptomes. PLoS ONE 2019, 14, e0219207. [Google Scholar] [CrossRef] [Green Version]
- Abdoulaye, A.H.; Foda, M.F.; Kotta-Loizou, I. Viruses infecting the plant pathogenic fungus Rhizoctonia solani. Viruses 2019, 11, 1113. [Google Scholar] [CrossRef] [Green Version]
- Owashi, Y.; Aihara, M.; Moriyama, H.; Arie, T.; Teraoka, T.; Komatsu, K. Population structure of double-stranded RNA mycoviruses that infect the rice blast fungus Magnaporthe oryzae in Japan. Front. Microbiol. 2020, 11, 593784. [Google Scholar] [CrossRef] [PubMed]
- Al-Rwahnih, M.; Daubert, S.; Urbez-Torres, J.R.; Cordero, F.; Rowhani, A. Deep sequencing evidence from single grapevine plants reveals a virome dominated by mycoviruses. Arch. Virol. 2011, 156, 397–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vainio, E.J.; Jurvansuu, J.; Streng, J.; Rajamaki, M.L.; Hantula, J.; Valkonen, J.P.T. Diagnosis and discovery of fungal viruses using deep sequencing of small RNAs. J. Gen. Virol. 2015, 96, 714–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thapa, V.; Roossinck, M.J. Determinants of coinfection in the mycoviruses. Front. Cell. Infect. Microbiol. 2019, 9, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, R.; van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 804. [Google Scholar] [CrossRef] [Green Version]
- Hirrel, M.C.; Lee, F.N.; Dale, J.L.; Pluntett, D.E. First report of sheath blight (Rhizoctonia solani) on field corn in Arkansas. Plant Dis. 1988, 72, 644. [Google Scholar] [CrossRef]
- Brown, M.; Jayaweera, D.P.; Hunt, A.; Woodhall, J.W.; Ray, R.V. Yield losses and control by sedaxane and fludioxonil of soilborne Rhizoctonia, Microdochium and Fusarium species in winter wheat. Plant Dis. 2021, 105, 2521–2530. [Google Scholar] [CrossRef]
- Yang, Y.G.; Zhao, C.; Guo, Z.J.; Wu, X.H. Anastomosis group and pathogenicity of Rhizoctonia solani associated with stem canker and black scurf of potato in China. Eur. J. Plant Pathol. 2015, 143, 99–111. [Google Scholar] [CrossRef]
- Balali, G.R.; Neate, S.M.; Scott, E.S.; Whisson, D.L.; Wicks, T.J. Anastomosis group and pathogenicity of isolates of Rhizoctonia solani from potato crops in South Australia. Plant Pathol. 1995, 44, 1050–1057. [Google Scholar] [CrossRef]
- Carling, D.E.; Brainard, K.A.; Virgen-Calleros, G.; Olalde-Portugal, V. First report of Rhizoctonia solani AG-7 on potato in Mexico. Plant Dis. 1998, 82, 127. [Google Scholar] [CrossRef]
- Woodhall, J.W.; Lees, A.K.; Edwards, S.G.; Jenkinson, P. Characterization of Rhizoctonia solani from potato in Great Britain. Plant Pathol. 2007, 56, 286–295. [Google Scholar] [CrossRef]
- Yanar, Y.; Yllmaz, G.; Cesmeli, I.; Coskun, S. Characterization of Rhizoctonia solani isolates from potatoes in Turkey and screening potato cultivars for resistance to AG-3 isolates. Phytoparasitica 2005, 33, 370–376. [Google Scholar] [CrossRef]
- Yang, Y.G.; Zhao, C.; Guo, Z.J.; Wu, X.H. Potato stem canker caused by binucleate Rhizoctonia AG-G in China. J Gen. Plant Pathol. 2015, 81, 287–290. [Google Scholar] [CrossRef]
- Das, S.; Shah, F.A.; Butler, R.C.; Falloon, R.E.; Stewart, A.; Raikar, S.; Pitman, A.R. Genetic variability and pathogenicity of Rhizoctonia solani associated with black scurf of potato in New Zealand. Plant Pathol. 2014, 63, 651–666. [Google Scholar] [CrossRef]
- Woodhall, J.W.; Adams, I.P.; Peters, J.C.; Harper, G.; Boonham, N. A new quantitative real-time PCR assay for Rhizoctonia solani AG-3 PT and the detection of AGs of Rhizoctonia solani associated with potato in soil and tuber samples in Great Britain. Eur. J. Plant Pathol. 2013, 136, 273–280. [Google Scholar] [CrossRef]
- Bandy, B.P.; Leach, S.S.; Tavantzis, S.M. Anastomosis group 3 is the major cause of Rhizoctonia disease of potato in Maine. Plant Dis. 1988, 72, 596–598. [Google Scholar] [CrossRef]
- Castanho, B.; Butler, E.E.; Shepherd, R.J. The association of double-stranded RNA with Rhizoctonia decline. Phytopathology 1978, 68, 1515–1519. [Google Scholar] [CrossRef] [Green Version]
- Andika, I.B.; Wei, S.; Cao, C.M.; Salaipeth, L.; Kondo, H.; Sun, L.Y. Phytopathogenic fungus hosts a plant virus: A naturally occurring cross-kingdom viral infection. Proc. Natl. Acad. Sci. USA 2017, 114, 12267–12272. [Google Scholar] [CrossRef] [Green Version]
- Zhong, J.; Chen, C.Y.; Gao, B.D. Genome sequence of a novel mycovirus of Rhizoctonia solani, a plant pathogenic fungus. Virus Genes 2015, 51, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Liu, C.; Zhang, M.L.; Yang, M.; Zhou, E.X. Diversity of dsRNA viruses infecting rice sheath blight fungus Rhizoctonia solani AG-1 IA. Rice Sci. 2018, 25, 57–60. [Google Scholar] [CrossRef]
- Bartholomäus, A.; Wibberg, D.; Winkler, A.; Pühler, A.; Schlüter, A.; Varrelmann, M. Identification of a novel mycovirus isolated from Rhizoctonia solani (AG 2-2 IV) provides further information about genome plasticity within the order Tymovirales. Arch. Virol. 2017, 162, 555–559. [Google Scholar] [CrossRef]
- Das, S.; Falloon, R.E.; Stewart, A.; Pitman, A.R. Novel mitoviruses in Rhizoctonia solani AG-3 PT infecting potato. Fungal Biol. 2016, 120, 338–350. [Google Scholar] [CrossRef]
- Strauss, E.E.; Lakshman, D.K.; Tavantzis, S.M. Molecular characterization of the genome of a partitivirus from the basidiomycete Rhizoctonia solani. J. Gen. Virol. 2000, 81, 549–555. [Google Scholar] [CrossRef]
- Zheng, L.; Zhang, M.L.; Chen, Q.G.; Zhu, M.H.; Zhou, E.X. A novel mycovirus closely related to viruses in the genus Alphapartitivirus confers hypovirulence in the phytopathogenic fungus Rhizoctonia solani. Virology 2014, 456, 220–226. [Google Scholar] [CrossRef] [Green Version]
- Picarelli, M.A.S.C.; Forgia, M.; Rivas, E.B.; Nerva, L.; Chiapello, M.; Turina, M.; Colariccio, A. Extreme diversity of mycoviruses present in isolates of Rhizoctonia solani AG2-2 LP from Zoysia japonica from Brazil. Front. Cell. Infect. Microbiol. 2019, 9, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.W.; Li, Y.T.; Hu, C.H.; Han, C.G.; Zhou, T.; Zhao, C.; Wu, X.H. Full genome sequence of a new mitovirus from the phytopathogenic fungus Rhizoctonia solani. Arch. Virol. 2020, 165, 1719–1723. [Google Scholar] [CrossRef] [PubMed]
- Bartholomäus, A.; Wibberg, D.; Winkler, A.; Puhler, A.; Schluter, A.; Varrelmann, M. Deep sequencing analysis reveals the mycoviral diversity of the virome of an avirulent isolate of Rhizoctonia solani AG-2-2 IV. PLoS ONE 2016, 11, e0165965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Gai, X.T.; Chen, R.X.; Li, C.X.; Zhao, G.K.; Xia, Z.Y.; Zou, C.M.; Zhong, J. Characterization of three novel betapartitiviruses co-infecting the phytopathogenic fungus Rhizoctonia solani. Virus Res. 2019, 270, 197649. [Google Scholar] [CrossRef] [PubMed]
- Lyu, R.L.; Zhang, Y.; Tang, Q.; Li, Y.Y.; Cheng, J.S.; Fu, Y.P.; Chen, T.; Jiang, D.H.; Xie, J.T. Two alphapartitiviruses co-infecting a single isolate of the plant pathogenic fungus Rhizoctonia solani. Arch. Virol. 2018, 163, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Morris, T.J.; Dodds, J.A. Isolation and analysis of double-stranded RNA from virus-infected plant and fungal tissue. Phytopathology 1979, 69, 854–858. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.T.; Ghabrial, S.A. Molecular characterizations of two mitoviruses co-infecting a hypovirulent isolate of the plant pathogenic fungus Sclerotinia sclerotiorum. Virology 2012, 428, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.Y.; Wu, S.S.; Liu, L.J.; Cheng, J.S.; Fu, Y.P.; Jiang, D.H.; Xie, J.T. A mitovirus related to plant mitochondrial gene confers hypovirulence on the phytopathogenic fungus Sclerotinia sclerotiorum. Virus Res. 2015, 197, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, Y.; Abraham, A.; Uesaka, K.; Kondo, H.; Suga, H.; Suzuki, N.; Chiba, S. Novel mitoviruses and a unique Tymo-like virus in hypovirulent and virulent strains of the Fusarium head blight fungus, Fusarium boothii. Viruses 2018, 10, 584. [Google Scholar] [CrossRef] [Green Version]
- Nibert, M.L.; Vong, M.; Fugate, K.K.; Debat, H.J. Evidence for contemporary plant mitoviruses. Virology 2018, 518, 14–24. [Google Scholar] [CrossRef]
- Stielow, J.B.; Bratek, Z.; Klenk, H.P.; Winter, S.; Menzel, W. A novel mitovirus from the hypogeous ectomycorrhizal fungus Tuber excavatum. Arch. Virol. 2012, 157, 787–790. [Google Scholar] [CrossRef]
- Herrero, N.; Zabalgogeazcoa, I. Mycoviruses infecting the endophytic and entomopathogenic fungus Tolypocladium cylindrosporum. Virus Res. 2011, 160, 409–413. [Google Scholar] [CrossRef] [Green Version]
- Ran, H.C.; Liu, L.J.; Li, B.; Cheng, J.S.; Fu, Y.P.; Jiang, D.H.; Xie, J.T. Co-infection of a hypovirulent isolate of Sclerotinia sclerotiorum with a new botybirnavirus and a strain of a mitovirus. Virol. J. 2016, 13, 92. [Google Scholar] [CrossRef] [Green Version]
- Hao, F.M.; Ding, T.; Wu, M.D.; Zhang, J.; Yang, L.; Chen, W.D.; Li, G.D. Two novel hypovirulence-associated mycoviruses in the phytopathogenic fungus Botrytis cinerea: Molecular characterization and suppression of infection cushion formation. Viruses 2018, 10, 254. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Liu, R.; Li, C.X.; Wang, H.; Zhu, H.J.; Gao, B.D.; Zhou, Q.; Zhong, J. A victorivirus and two novel mitoviruses co-infected the plant pathogen Nigrospora oryzae. Viruses 2019, 11, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hisano, S.; Zhang, R.; Faruk, M.I.; Kondo, H.; Suzuki, N. A neo-virus lifestyle exhibited by a (+) ssRNA virus hosted in an unrelated dsRNA virus: Taxonomic and evolutionary considerations. Virus Res. 2018, 244, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Hisano, S.; Tani, A.; Kondo, H.; Kanematsu, S.; Suzuki, N. A capsidless ssRNA virus hosted by an unrelated dsRNA virus. Nat. Microbiol. 2016, 1, 15001. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Liu, S.X.; Chiba, S.; Kondo, H.; Kanematsu, S.; Suzuki, N. A novel single-stranded RNA virus isolated from a phytopathogenic filamentous fungus, Rosellinia necatrix, with similarity to hypo-like viruses. Front. Microbiol. 2014, 5, 360. [Google Scholar] [CrossRef]
- Kwon, S.J.; Lim, W.S.; Park, S.H.; Park, M.R.; Kim, K.H. Molecular characterization of a dsRNA mycovirus, Fusarium graminearum virus-DK21, which is phylogenetically related to hypoviruses but has a genome organization and gene expression strategy resembling those of plant potex-like viruses. Mol. Cells 2009, 23, 304–315. [Google Scholar] [CrossRef]
- Honda, S.; Eusebio-Cope, A.; Miyashita, S.; Yokoyama, A.; Aulia, A.; Shahi, S.; Kondo, H.; Suzuki, N. Establishment of Neurospora crassa as a model organism for fungal virology. Nat. Commum. 2020, 11, 2567. [Google Scholar] [CrossRef]
- Son, M.; Lee, K.M.; Yu, J.; Kang, M.; Park, J.M.; Kwon, S.J.; Kim, K.H. The HEX1 gene of Fusarium graminearum is required for fungal asexual reproduction and pathogenesis and for efficient viral RNA accumulation of Fusarium graminearum virus 1. J. Virol. 2013, 87, 10356–10367. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Lee, K.M.; Cho, W.K.; Park, J.Y.; Kim, K.H. Differential contribution of RNA interference components in response to distinct Fusarium graminearum virus infections. J. Virol. 2018, 92, e01756-17. [Google Scholar] [CrossRef] [Green Version]
- Nerva, L.; Vigani, G.; di Silvestro, D.; Ciuffo, M.; Forgia, M.; Chitarra, W.; Turina, M. Biological and molecular characterization of Chenopodium quinoa mitovirus 1 reveals a distinct small RNA response compared to those of cytoplasmic RNA viruses. J. Virol. 2019, 93, e01998-18. [Google Scholar] [CrossRef] [Green Version]
- Paquin, B.; Laforest, M.J.; Forget, L.; Roewer, I.; Wang, Z.; Longcore, J.; Lang, B.F. The fungal mitochondrial genome project: Evolution of fungal mitochondrial genomes and their gene expression. Curr. Genet. 1997, 31, 380–395. [Google Scholar] [CrossRef]
- Nibert, M.L. Mitovirus UGA (Trp) codon usage parallels that of host mitochondria. Virology 2017, 507, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Li, W.; Chen, H.G.; Yu, H.S. Full genome sequence of a putative novel mitovirus isolated from Rhizoctonia cerealis. Arch. Virol. 2015, 160, 1815–1818. [Google Scholar] [CrossRef] [PubMed]
- Abdoulaye, A.H.; Cheng, J.S.; Fu, Y.P.; Jiang, D.H.; Xie, J.T. Complete genome sequence of a novel mitovirus from the phytopathogenic fungus Rhizoctonia oryzae-sativae. Arch. Virol. 2017, 162, 1409–1412. [Google Scholar] [CrossRef] [PubMed]
- Doherty, M.; Coutt, R.H.A.; Brasier, C.M.; Buck, K.W. Sequence of RNA-dependent RNA polymerase genes provides evidence for three more distinct mitoviruses in Ophiostoma novo-ulmi isolate Ld. Virus Genes 2006, 33, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.G.; Cole, T.E.; Brasier, C.M.; Buck, K.W. Evolutionary relationships among putative RNA-dependent RNA polymerases encoded by a mitochondrial virus-like RNA in the Dutch elm disease fungus, Ophiostoma novo-ulmi, by other viruses and virus-like RNAs and by the Arabidopsis mitochondrial genome. Virology 1998, 246, 158–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalifa, M.E.; Pearson, M.N. Molecular characterization of novel mitoviruses associated with Sclerotinia sclerotiorum. Arch. Virol. 2014, 159, 3157–3160. [Google Scholar] [CrossRef]
- Khalifa, M.E.; Pearson, M.N. Molecular characterization of three mitoviruses co-infecting a hypovirulent isolate of Sclerotinia sclerotiorum fungus. Virology 2013, 441, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Liu, M.; Zhu, H.J.; Zhong, J.; Liao, X.L.; Zhou, Q. Molecular characterization of a novel mitovirus from the plant pathogenic fungus Botryosphaeria dothidea. Arch. Virol. 2021, 166, 633–637. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Wang, J.X.; Yang, B.; Wang, Q.R.; Zhou, J.J.; Yu, W.F. Molecular characterization of a debilitation-associated partitivirus infecting the pathogenic fungus Aspergillus flavus. Front. Microbiol. 2019, 10, 626. [Google Scholar] [CrossRef]
- Zhong, J.; Chen, D.; Lei, X.H.; Zhu, H.J.; Zhu, J.Z.; Gao, B.D. Detection and characterization of a novel gammapartitivirus in the phytopathogenic fungus Colletotrichum acutatum strain HNZJ001. Virus Res. 2014, 190, 104–109. [Google Scholar] [CrossRef]
- Hong, Y.G.; Dover, S.L.; Cole, T.E.; Brasier, C.M.; Buck, K.W. Multiple mitochondrial viruses in an isolate of the Dutch elm disease fungus Ophiostoma novo-ulmi. Virology 1999, 258, 118–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nerva, L.; Silvestri, A.; Ciuffo, M.; Palmano, S.; Varese, G.C.; Turina, M. Transmission of Penicillium aurantiogriseum partiti-like virus 1 to a new fungal host (Cryphonectria parasitica) confers higher resistance to salinity and reveals adaptive genomic changes. Environ. Microbiol. 2017, 19, 4480–4492. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Liu, H.Q.; Zhang, M.L.; Cao, X.; Zhou, E.X. The complete genomic sequence of a novel mycovirus from Rhizoctonia solani AG-1 IA strain B275. Arch. Virol. 2013, 158, 1609–1612. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Xu, P.; Zhang, L.F.; Xia, Z.Y.; Qin, X.Y.; Yang, G.H.; Mo, X.H. Molecular characterization of a novel mycovirus from Rhizoctonia fumigata AG-Ba isolate C-314 Baishi. Arch. Virol. 2015, 160, 2371–2374. [Google Scholar] [CrossRef]
- Liu, C.C.; Li, M.; Redda, E.T.; Mei, J.; Zhang, J.T.; Elena, S.F.; Wu, B.L.; Jiang, X.L. Complete nucleotide sequence of a novel mycovirus from Trichoderma harzianum in China. Arch. Virol. 2019, 164, 1213–1216. [Google Scholar] [CrossRef] [Green Version]
- Chiba, S.; Salaipeth, L.; Lin, Y.H.; Sasaki, A.; Kanematsu, S.; Suzuki, N. A novel bipartite double-stranded RNA mycovirus from the white root rot fungus Rosellinia necatrix: Molecular and biological characterization, taxonomic considerations, and potential for biological control. J. Virol. 2009, 83, 12801–12812. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.D.; Jin, F.Y.; Zhang, J.; Yang, L.; Jiang, D.H.; Li, G.Q. Characterization of a novel bipartite double-stranded RNA mycovirus conferring hypovirulence in the phytopathogenic fungus Botrytis porri. J. Virol. 2012, 86, 6605–6619. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.H.; Wang, Y.; Sun, X.Z.; Cheng, J.S.; Fu, Y.P.; Liu, H.Q.; Jiang, D.H.; Ghabrial, S.A.; Xie, J.T. Characterization of a novel megabirnavirus from Sclerotinia sclerotiorum reveals horizontal gene transfer from single-stranded RNA virus to double-stranded RNA virus. J. Virol. 2015, 89, 8567–8579. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.F.; Zhao, H.; Xue, C.Y.; Xu, C.; Geng, Y.H.; Zang, R.; Guo, Y.X.; Wu, H.Y.; Zhang, M. Complete genome sequence of a novel mycovirus isolated from the phytopathogenic fungus Corynespora cassiicola in China. Arch. Virol. 2020, 165, 2401–2404. [Google Scholar] [CrossRef]
- Liu, H.; Wang, H.; Zhou, Q. A novel mycovirus isolated from the plant pathogenic fungus Botryosphaeria dothidea. Arch. Virol. 2021, 166, 1267–1272. [Google Scholar] [CrossRef]
- Yu, J.; Kwon, S.J.; Lee, K.M.; Son, M.; Kim, K.H. Complete nucleotide sequence of double-stranded RNA viruses from Fusarium graminearum strain DK3. Arch. Virol. 2009, 154, 1855–1858. [Google Scholar] [CrossRef] [PubMed]
- Vainio, E.J.; Hyder, R.; Aday, G.; Hansen, E.; Piri, T.; Dogmus-Lehtijarvi, T.; Lehtijarvi, A.; Korhonen, K.; Hantula, J. Population structure of a novel putative mycovirus infecting the conifer root-rot fungus Heterobasidion annosum sensu lato. Virology 2012, 422, 366–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koonin, E.V.; Dolja, V.V. Evolution and taxonomy of positive-strand RNA viruses: Implications of comparative analysis of amino acid sequences. Crit. Rev. Biochem. Mol. 1993, 28, 375–430. [Google Scholar] [CrossRef] [PubMed]
- Ahlquist, P. Parallels among positive-strand RNA viruses, reverse-transcribing viruses and double-stranded RNA viruses. Nat. Rev. Microbiol. 2006, 4, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Wolf, Y.I.; Kazlauskas, D.; Iranzo, J.; Lucia-Sanz, A.; Kuhn, J.H.; Krupovic, M.; Dolja, V.V.; Koonin, E.V. Origins and evolution of the global RNA virome. mBio 2018, 9, e02329-18. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Name | Accession Number | Contig ID | Contig Length (nt) | Full Length (nt) | BLASTx First Hit | Identity | Query Cover |
|---|---|---|---|---|---|---|---|
| Rhizoctonia solani fusarivirus 4 (RsFV4) | MZ636366 | contig978 | 10,416 | 10,541 | Rhizoctonia solani fusarivirus 1 | 49.21% | 32% |
| Rhizoctonia solani fusarivirus 5 (RsFV5) | MW713065 | contig6043 | 8134 | >8140 | Rhizoctonia solani fusarivirus 1 | 71.34% | 49% |
| Rhizoctonia solani mitovirus 40 (RsMV40) | MW582547 | First_contig5 | 2806 | 3037 | Epicoccum nigrum mitovirus 1 | 73.97% | 81% |
| Rhizoctonia solani partitivirus 10 (RsPV10) | MW523007 | contig3112 | 2001 | 2029 | White clover cryptic virus 1 | 61.33% | 86% |
| MZ467297 | contig1221 | 1700 | 1713 | White clover cryptic virus 1 | 29.44% | 30% | |
| Rhizoctonia solani partitivirus 11 (RsPV11) | MW523009 | contig644 | 1921 | 1946 | Ceratobasidium partitivirus | 55.65% | 89% |
| MW523010 | contig315 | 1788 | 1833 | Ceratobasidium partitivirus | 24.49% | 82% | |
| Rhizoctonia solani RNA virus 11 (RsRV11) | MT648469 | contig14243 | 1730 | 1736 | Rhizoctonia fumigata mycovirus | 72.40% | 91% |
| MT648470 | contig11433 | 2300 | 2311 | Rhizoctonia fumigata mycovirus | 56.13% | 79% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Li, S.; Zhao, Y.; Zhou, T.; Wu, X.; Zhao, C. Six Novel Mycoviruses Containing Positive Single-Stranded RNA and Double-Stranded RNA Genomes Co-Infect a Single Strain of the Rhizoctonia solani AG-3 PT. Viruses 2022, 14, 813. https://doi.org/10.3390/v14040813
Li Y, Li S, Zhao Y, Zhou T, Wu X, Zhao C. Six Novel Mycoviruses Containing Positive Single-Stranded RNA and Double-Stranded RNA Genomes Co-Infect a Single Strain of the Rhizoctonia solani AG-3 PT. Viruses. 2022; 14(4):813. https://doi.org/10.3390/v14040813
Chicago/Turabian StyleLi, Yuting, Siwei Li, Yumeng Zhao, Tao Zhou, Xuehong Wu, and Can Zhao. 2022. "Six Novel Mycoviruses Containing Positive Single-Stranded RNA and Double-Stranded RNA Genomes Co-Infect a Single Strain of the Rhizoctonia solani AG-3 PT" Viruses 14, no. 4: 813. https://doi.org/10.3390/v14040813